

Introduction
Les marais du Parc naturel régional (Pnr) de Brière jouent un rôle fondamental dans la préservation de la biodiversité. Ils sont le lieu de vie de nombreuses espèces végétales et animales paludicoles (Eybert et al, 2007 ; Clément et Francez, 2007), dont beaucoup sont menacées et protégées. Leur importance est notamment soulignée par l’existence de plusieurs périmètres de conservation définis par la législation Natura 2000 (PnrB, 2003 ; PnrB, 2007). Dans ce contexte, accroître la connaissance sur ces zones humides s’avère essentiel.
Pour répondre à cet objectif, ce travail propose une mise à jour de la cartographie fine de ces marais, d’autant plus indispensable qu’il n’existe que deux cartes des groupements végétaux de l’ensemble des zones humides du Parc. La plus ancienne (Clément et al., 1980), bien que précise, ne prend pas en compte l’évolution de la végétation de ces trente dernières années. La plus récente (Geffray, 2001) est fondée sur l’analyse d’images SPOT de moyenne résolution spatiale, ce qui ne lui permet pas d’atteindre une précision convenable pour cartographier les habitats. Pour pallier ces manques, nous avons choisi de tester la méthode de cartographie par analyse orientée-objet sur des images de télédétection à Très Haute Résolution Spatiale (THRS).
À terme, la cartographie produite a pour but de contribuer à la définition des habitats potentiels de deux espèces sensibles d’oiseaux paludicoles, particulièrement représentées sur le territoire du Pnr : le Phragmite aquatique (Acrocephalus paludicola) (Marquet et Séchet, 2010), et le Butor étoilé (Botaurus stellaris) (Marquet, 2014). Nous nous sommes donc intéressés aux habitats privilégiés de ces espèces, en période migratoire pour le Phragmite aquatique (Le Nevé et al., 2009), ou en reproduction pour le Butor étoilé (Hunault et Kerbiriou, 2007). Il s’agit des groupements végétaux dominés par les grandes hélophytes, plantes dont l’appareil racinaire reste toute l’année dans les vases humides (notamment les roseaux Phragmites australis et Phalaris arundinacea, les laîches Carex spp., et les joncs Scirpus spp. ou Eleocharis spp.).
La réalisation de ce travail a été permise par un partenariat entre le Parc naturel régional de Brière et le laboratoire LETG-Nantes Géolittomer (UMR 6554), et grâce au soutien du programme ISIS-Pléiades du CNES.
1. La végétation de zones humides difficile d’accès
Une marqueterie de formes
Les zones humides du Pnr de Brière suivent les lits de deux cours d’eau (fig. 1) : le Brivet, dernier affluent de la Loire, associé aux marais de Brière et de Donges, et le Mès, fleuve côtier situé à l’ouest de la Brière, caractérisé dans sa partie maritime par l’exploitation de marais salants. Les différentes formes de végétation y sont dépendantes des hauteurs d’eau, elles-mêmes conditionnées par les variations topographiques. Cependant, elles sont aussi tributaires d’autres facteurs, comme les pratiques agricoles, ou encore les caractéristiques du sol.
Figure 1 – localisation de la zone d’étude
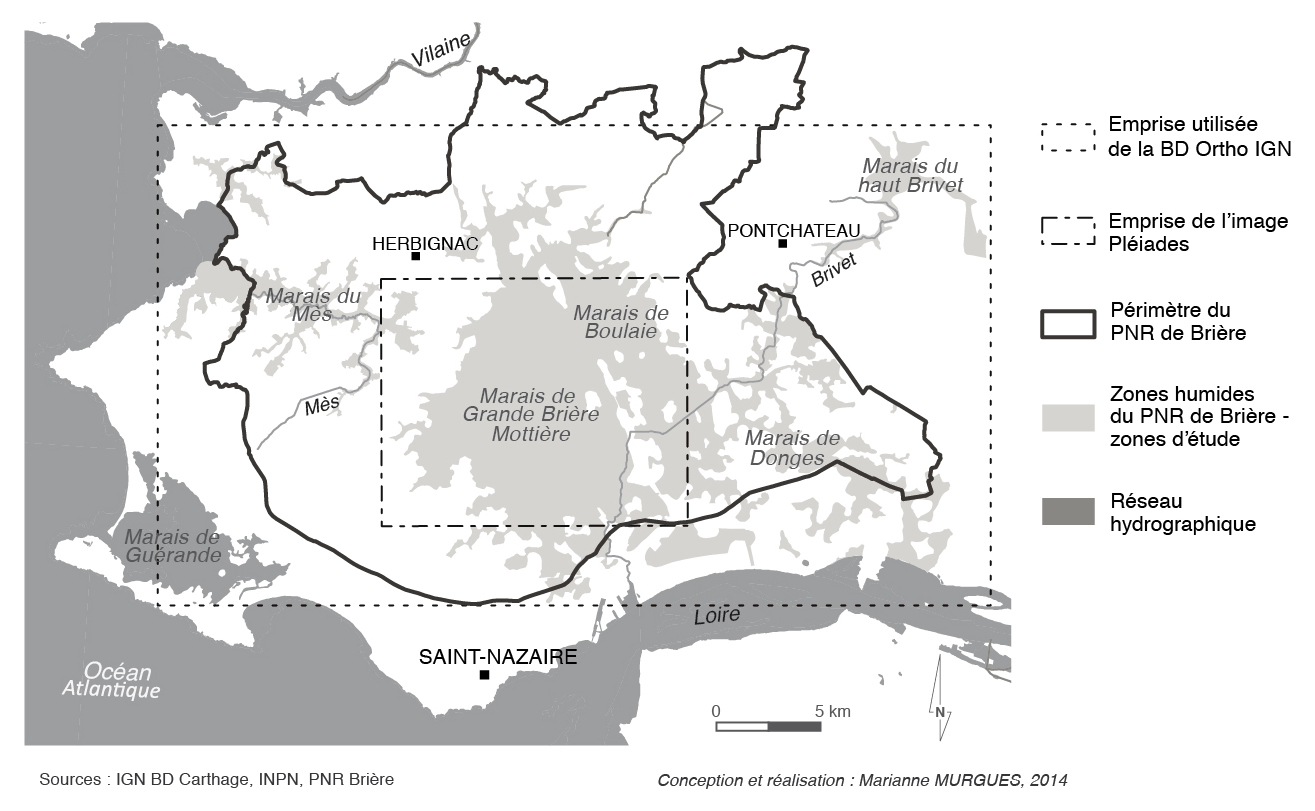
Plusieurs types de végétation caractérisent ces espaces, en fonction du degré d’hygrophilie (tolérance à l’eau) et d’halophilie (tolérance au sel) des différentes espèces. Certaines formes, comme les roselières (groupements dominés par de grandes hélophytes), sont très représentées dans les zones supportant une longue période d’inondation (environ dix mois). Les formations prairiales, inondées sur une période généralement plus courte (un à cinq mois), marquent elles aussi largement les paysages de ces zones humides. D’autres formations végétales ponctuent ces espaces, comme les herbiers aquatiques, les boisements humides ou les gazons amphibies. L’ensemble des groupements végétaux forme ainsi un patron complexe, structuré et étagé par la durée d’inondation et le niveau de l’eau.
Une évolution constante
La composition et la dynamique des peuplements végétaux des zones humides de Brière s’expliquent en grande partie par les variations interannuelles des niveaux d’eau. Cependant, deux autres facteurs peuvent en expliquer les changements rapides. En premier lieu, le déclin des pratiques agricoles favorise des dynamiques d’atterrissement et de fermeture des habitats, par le développement en bordure de marais et au sein de certaines roselières de fourrés et boisements humides. En second lieu, la pression grandissante d’espèces exotiques a pu modifier certains paysages et habitats au cours des deux dernières décennies, notamment en Grande Brière Mottière, territoire emblématique au sein de ces zones humides. A titre d’exemple, le développement d’herbiers denses de Jussie (Ludwigia sp.) change l’aspect des canaux, vases nues et prairies hygrophiles. Les herbiers aquatiques de Nénuphar blanc (Nymphea alba), autrefois si caractéristiques des plans d’eau briérons (Clément et al., 1980), ont quant à eux disparu, en concomitance avec l’introduction dans le marais de l’écrevisse de Louisiane (Procambarus clarkii). Une mise à jour fréquente de la cartographie semble donc aujourd’hui plus que jamais d’une grande importance pour le suivi et la compréhension de ces évolutions.
Un accès difficile
La méthode la plus précise et la plus fréquente pour produire une carte de la végétation est l’approche botanique de terrain. Cependant, à l’échelle d’un marais de 25 000 ha, elle est inadaptée. En effet, si la taille de la surface à couvrir est déjà une difficulté, la nature même du terrain augmente la contrainte. La plupart des déplacements de longue distance s’effectuent en barque, et les déplacements au sein des roselières sont rendus difficiles par la vase, la végétation haute et dense, et la présence de l’eau. La méthode de cartographie choisie ici permet d’éviter ces difficultés.
2. Des outils et méthodes adaptés : images THRS et analyse orientée-objet
Les différentes étapes de ce protocole et leur succession sont résumées dans la figure 2.
Figure 2 – description du protocole méthodologique appliqué pour la cartographie de la végétation
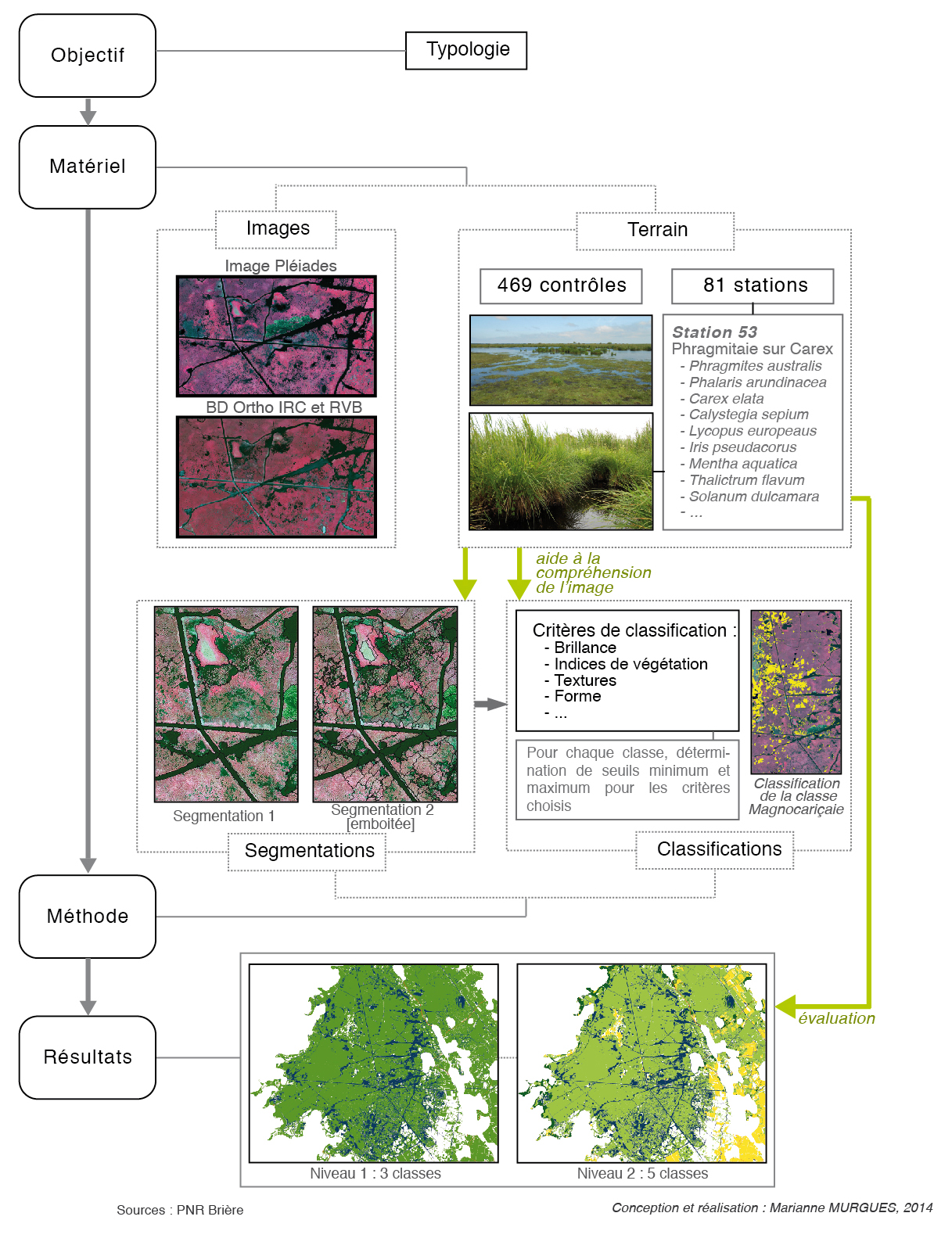
L’originalité de cette méthode tient avant tout au type d’image employée et au type d’analyse menée : image THRS et analyse orientée-objet. Néanmoins, elle repose aussi sur d’autres éléments plus classiques en cartographie : la typologie et le terrain.
La typologie
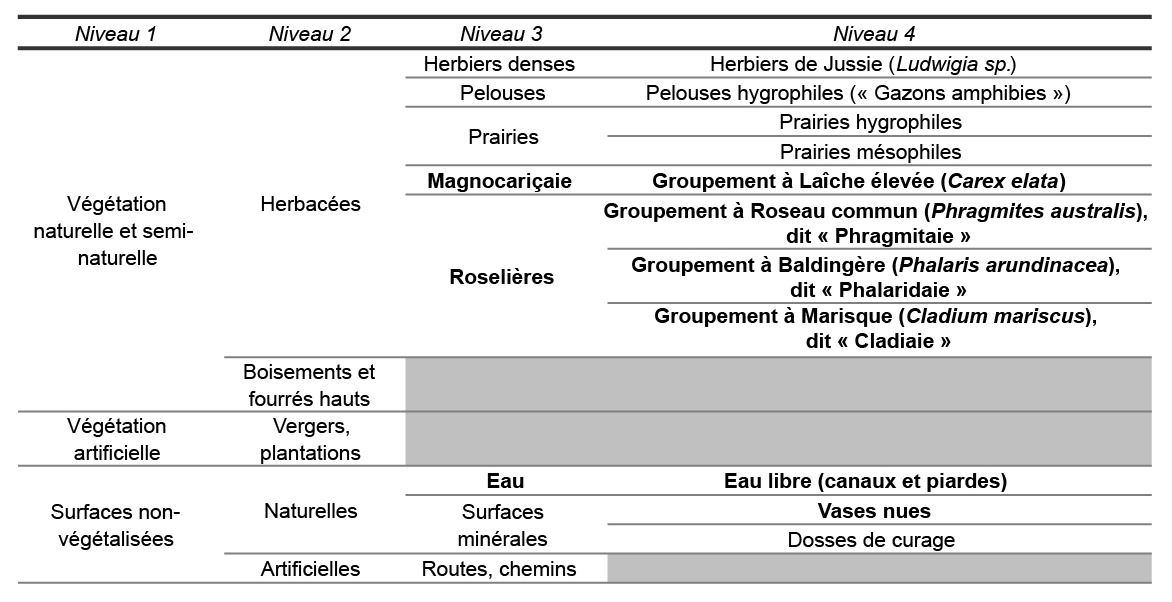
La typologie retenue s’appuie sur les référentiels en matière de classification des habitats, CORINE Biotopes notamment (Bissardon et al., 1997), mais aussi sur une classification emboîtée de la végétation proposée par le Conservatoire botanique national de Brest (Delassus et Magnanon, 2013), et enfin sur des travaux menés sur la Brière (Murgues, 2013). Elle présente quatre niveaux emboîtés (tab. 1).
Tableau 1 – typologie de travail (non-exhaustive du point de vue phytosociologique) pour la cartographie des végétations des zones humides du Pnr de Brière. Les classes en gras sont les classes prioritaires pour cette étude.
Le matériel : des images à Très Haute Résolution Spatiale (THRS)
Les images de télédétection, aériennes ou spatiales, rendent possible l’accès à l’information à distance et de façon continue sur les espaces observés. Compte tenu des objectifs de ce travail, nous avons utilisé des images THRS, dont les pixels représentent au sol un carré de moins d’un mètre de côté. Cette précision est rendue possible par les progrès récents en matière de résolution spatiale (Sellin et al., 2013). La question centrale de l’utilisation de ces sources d’information est celle de l’adéquation entre le niveau typologique souhaité, décrivant la végétation, et les caractéristiques de l’image en matière de résolution spatiale, spectrale et temporelle.
Les images aériennes, et notamment la BD Ortho® de l’IGN, se présentent à priori comme un support adapté pour cette cartographie. Elles offrent en effet une résolution spatiale très fine, qui contrebalance leur faible résolution spectrale : 30 cm pour les images RVB (canaux Bleu, Vert, Rouge), et 50 cm pour les images IRC (canaux Vert, Rouge et Proche Infra-Rouge). Ces ortho-images présentent néanmoins trois inconvénients majeurs pour la cartographie de la végétation. D’abord, il n’est pas possible d’en choisir la date d’acquisition (ici les 16 et 23 juin 2009 pour la couverture de l’ensemble du Pnr). Ensuite, la répétitivité des données est faible : tous les trois ans depuis 2009. Enfin, la qualité radiométrique de l’image est très hétérogène du fait de l’assemblage de prises de vues effectuées à des dates et heures différentes, ce qui a posé de nombreux problèmes lors des procédures de classification.
Nous avons donc choisi d’expérimenter l’analyse orientée-objet sur un autre type d’image, à la résolution spatiale comparable. Il s’agit d’images acquises par le satellite Pléiades. Leur résolution spatiale est de 2 m en mode multispectral (4 bandes : Bleu, Vert, Rouge et Proche Infra-rouge), et de 50 cm en mode panchromatique. L’acquisition de ces données satellitaires a fait l’objet d’une programmation en fin de période d’inondation (20 août 2013). Pour des raisons de conflit avec d’autres demandes, il n’a pas été possible d’enregistrer des données à cette date sur l’ensemble du territoire du Pnr de Brière. L’acquisition a donc été centrée sur le marais de Grande Brière Mottière (fig. 1), le plus concerné par les groupements de grandes hélophytes.
Cependant, afin d’étendre la cartographie à l’ensemble des marais du Pnr, les résultats obtenus à partir de l’image Pléiades (fig. 3) ont été complétés par une analyse des images de la BD Ortho® (fig. 4). Un « masque » a été appliqué à ces deux images, afin de limiter l’analyse aux zones humides du Pnr.
Le protocole méthodologique : l’analyse d’image orientée-objet
Le protocole méthodologique est fondé sur les principes de l’analyse d’images dite orientée-objet, ici réalisée grâce au logiciel eCognition 8.7TM (Trimble®). Du fait de sa complexité, ce protocole n’est pas ici intégralement détaillé. Il est synthétisé à travers les deux principes qui font son originalité et son efficacité : la segmentation et la classification.
La segmentation est le processus de « découpage » de l’image, qui crée des groupements de pixels (les « objets »). Il s’agit dans notre analyse d’une segmentation dite « multi-résolution » (Baatz et Schäpe, 2000), qui procède par regroupement de pixels homogènes (forme et couleur) à des niveaux de détail considérés comme pertinents par rapport aux niveaux de la typologie. Toute la difficulté est donc de déterminer ces niveaux de détail : une segmentation trop grossière regroupe les éléments à distinguer, tandis qu’une segmentation trop fine crée des entités qui n’en sont pas, et qui sont ensuite difficiles à classer. Pour la distinction de la classe « Magnocariçaies » par exemple, une segmentation grossière ne permet pas de distinguer les éléments de cette classe de ceux de la classe « Groupements mixtes roselières-magnocariçaies » ; une segmentation fine au contraire génère des polygones trop petits pour répondre ensuite aux critères de définition de la classe.
La classification quant à elle est le processus visant à regrouper les objets par classes, selon leurs caractéristiques. Elle est fondée sur les valeurs spectrales de l’image (moyenne des valeurs des pixels fusionnés au sein d’un même objet), mais aussi sur de nombreux autres critères discriminants comme la forme des objets, leur texture, leur localisation ou encore les relations hiérarchiques entre les classes. Là encore, il s’agit donc d’identifier et de choisir pour une classe donnée les critères permettant de la distinguer. A titre d’exemple, les canaux, remplis d’une eau chargée en matière organique, se différencient des autres types de surfaces par leur signature spectrale et leur couleur, mais aussi par leur forme (grande longueur et faible largeur). Cette recherche de critères adaptés s’opère pour chaque classe, jusqu’à ce que la classification soit acceptable.
L’approche complémentaire du terrain
Un travail d’identification sur le terrain a été effectué pour permettre la désignation des différents groupements végétaux, et notamment des groupements de grandes hélophytes, desquels il n’existait pratiquement aucun recensement. Au total, 200 points d’observation ont été réalisés dans les marais du Mès et du haut-Brivet, et 350 dans les marais de Brière, Boulaie et Donges au cours du printemps et de l’été 2013. Parmi ces derniers, 81 stations de relevés de végétation selon la méthode phytosociologique ont été échantillonnées. En zone humide, une telle approche demande beaucoup de temps du fait des contraintes d’accès précédemment citées ; elle n’a donc pas pu être menée de façon exhaustive.
L’évaluation des résultats
Le niveau de fiabilité des cartes obtenues (fig. 3 et 4) a pu être évalué en procédant à l’analyse d’une matrice de confusion entre la carte et la réalité du terrain. Cette dernière est représentée notamment par les 550 points d’observation précédemment cités, ainsi que par des données disponibles au Pnr : des inventaires botaniques effectués entre 2009 et 2012, et la cartographie linéaire des canaux. Le calcul de l’indice de Kappa (K) effectué à partir de cette matrice (Girard et Girard, 1999) permet de quantifier la correspondance entre carte et terrain. L’indice est compris entre les valeurs 0 (correspondance nulle) et 1 (correspondance parfaite). Un résultat au-dessus de 0,6 est considéré comme « bon » (id.). Dans le détail, on distingue la précision pour l’utilisateur (PU) de la précision pour le réalisateur (PR), exprimées en pourcentage. Nous donnerons ici préférentiellement la PU, qui indique combien les objets contenus dans une classe ont de chance d’identifier réellement au sol les entités correspondant à cette classe. À l’inverse, la PR indiquerait combien une entité « réelle » a de chances d’être effectivement attribuée à la classe correspondante.
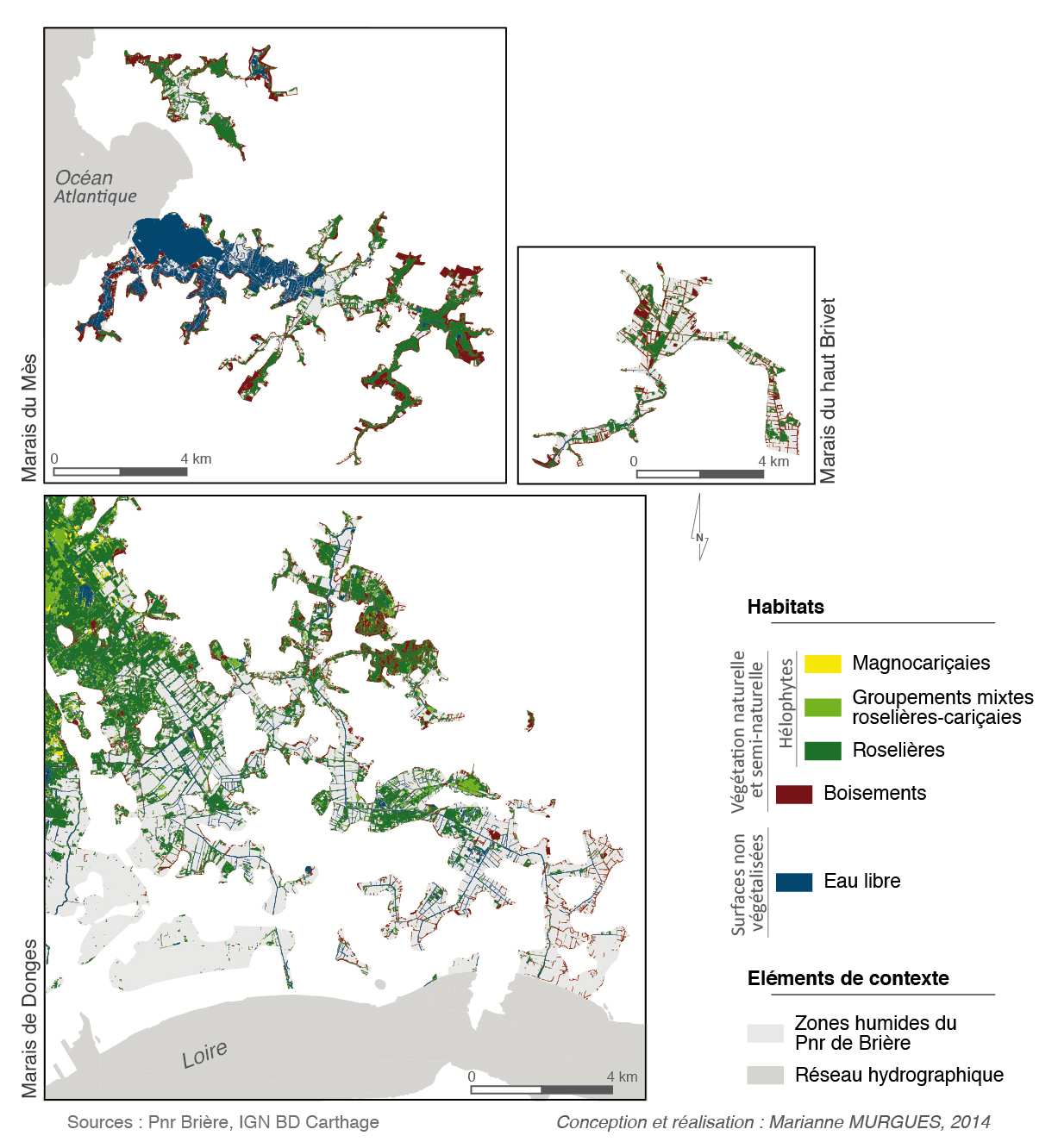
Figure 3 – répartition des habitats des zones humides des marais de Brière et de Boulaie en 2013, obtenue par analyse d’image orientée-objet, d’après une image Pléiades (20 août 2013). Cartographie réalisée en 2013.
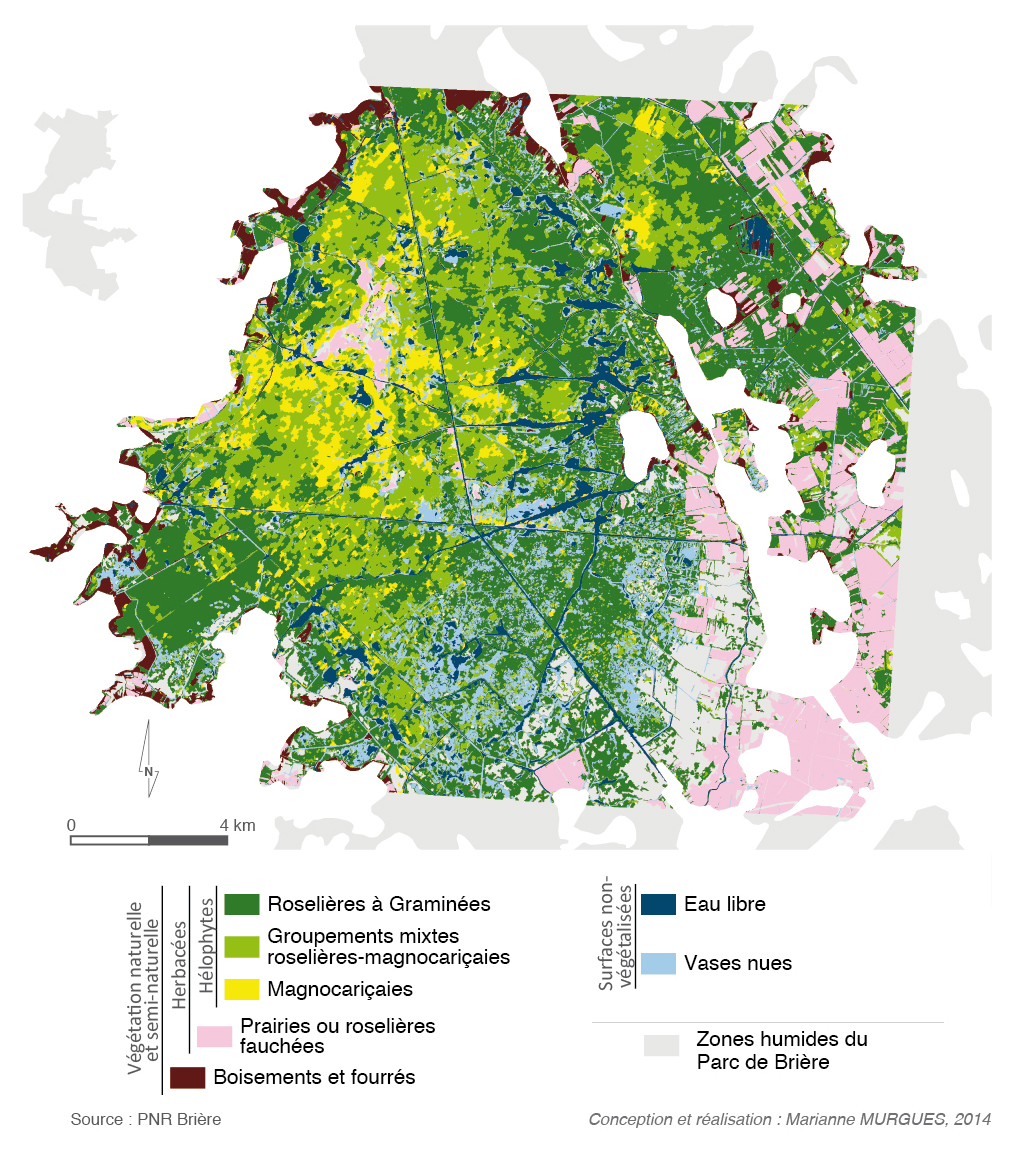
Figure 4 – répartition des habitats de grandes hélophytes, des boisements et de l’eau libre dans les zones humides du Pnr de Brière en 2009, obtenue par analyse d’image orientée-objet, d’après les BD Ortho® de l’IGN (16 et 23 juin 2009). Cartographie réalisée en 2013. Ces cartes, extraites d’une carte générale obtenue à partir des BD Ortho®, complètent celle obtenue par l’image Pléiades, en couvrant des espaces situés hors de son périmètre.
3. Des apports sur la connaissance et la conservation des zones humides
Analyse des résultats
Comme indiqué plus haut, l’ensemble du marais n’est pas cartographié avec la même source. Nous présentons donc ici d’une part la cartographie du marais de Grande Brière Mottière (fig. 3) obtenue à partir de l’image satellitaire Pléiades de 2013, et d’autre part celles des marais du Mès, du haut Brivet et de Donges (fig. 4), extraites de la carte générale obtenue à partir des ortho-images de 2009 (BD Ortho® de l’IGN).
Les légendes de ces cartes diffèrent légèrement de la typologie de travail (tab. 1). Cette typologie présente en effet les groupements qui devaient être distingués, tandis que la carte ne présente que les groupements qui ont pu être distingués. Il est important de retenir qu’il existe un décalage entre ce que l’œil humain perçoit, et ce qu’un algorithme identifie, certaines nuances échappant totalement à ce dernier. Ainsi, les sous-classes de roselières « Phragmitaie », « Phalaridaie » et « Cladiaie » (niveau 4 de la typologie) n’ont pu être distinguées de façon convenable, en dépit de différences de teintes et de textures perceptibles à l’œil. Au contraire, la sous-classe correspondant aux groupements mixtes roselière-magnocariçaie, se distinguant assez nettement des autres roselières comme des magnocariçaies, est devenue une classe à part entière ; cette distinction se justifie aussi par le faciès intermédiaire du groupement. Enfin, la classe « Boisements », groupement non-prioritaire ici, est présentée sur les deux cartes. De par leur forme et leur texture, les boisements se distinguaient en effet nettement des autres formes de végétation. Classer immédiatement ce groupement a permis de donner des repères paysagers utiles pour la suite des traitements, le voisinage étant un critère possible pour les classifications.
Dans le détail, la carte des marais de Brière et de Boulaie (fig. 3) (K = 0,76) montre une prédominance des formes de grandes hélophytes (PU = 67,2 %). La classe « magnocariçaies » (PU = 64,5 %) est relativement concentrée dans la partie nord-ouest du marais de Brière, et au nord des marais de la Boulaie. La classe « roselières » (PU = 73,4 %) se retrouve dans une large périphérie des deux marais. La classe « groupements mixtes roselière-magnocariçaie » (PU = 51,9 %) est relativement étendue et semble dépasser le simple rôle d’intermédiaire entre les deux premières classes. Ces trois classes sont sujettes à quelques confusions résiduelles entre elles, ainsi qu’avec certains groupements hygrophiles non cartographiés. Cette carte présente aussi de bons résultats pour les vases nues (PU = 93,8 %), qui occupent la périphérie des trous d’eau hivernaux et printaniers au sein des roselières (fig. 4) ; elles n’apparaissent pas dans les canaux principaux, qui restent inondés toute l’année (PU de l’eau libre = 97,9 %). Enfin, du fait de sa date, l’image Pléiades permet aussi de cartographier les surfaces fauchées. Celles-ci, assez différentes des vases par leur réflectance, et par leur forme relativement géométrique, ont été distinguées dans la classe « prairies et roselières fauchées » (PU = 93,3 %), qui n’apparaissait pas en typologie. Cette classe se retrouve en périphérie du marais le plus inondé (et donc des classes de roselières et de magnocariçaies), et sur la Butte des Pierres (partie nord-ouest du marais).
Les cartes des résultats pour les marais du Mès (K = 0,53) et du haut Brivet (K = 0,56) (fig. 4) ne présentent que trois classes au total, les groupements à Carex elata (classes « magnocariçaie » et « groupement mixte roselière-magnocariçaie ») ne s’observant pas dans ces deux marais. Dans les marais de Donges (K = 0,7), ces groupements s’observent aussi assez peu ; seuls les « groupements mixtes roselières-magnocariçaies » (PU = 57,7 %) sont visibles. De manière générale, les groupements de grandes hélophytes sont ici moins présents que dans les marais de Brière et de Boulaie. Les marais de Donges, du Mès et du haut Brivet sont en effet beaucoup plus anthropisés : les nombreux espaces non-classés correspondant à des prairies, ou à des constructions humaines. Il est intéressant d’observer l’apport de cette méthode dans la cartographie des réseaux d’eau. Dans les marais de Donges (PU = 98,2 %) comme du haut Brivet (PU = 86,7 %), on observe ainsi nettement les canaux de drainage agricoles, et dans les marais du Mès (PU = 93,6 %), on distingue les œillets des marais salants. Dans ces trois marais, la classe des boisements présente là encore des résultats satisfaisants (PU Donges = 84,3 % ; PU Mès = 78,6 % ; PU haut Brivet = 71,7 %).
Cartographie et suivi de groupements végétaux
Au-delà de la précision spatiale, la précision sémantique atteinte est appréciable. En effet, les cartes produites distinguent entre eux différents groupements de grandes hélophytes. Ce niveau de détail n’avait pu être atteint dans le travail précédemment cité de Geffray (2001), reposant sur une méthode d’analyse plus classique (dite « pixel ») et des images SPOT. Celles-ci avaient une résolution spectrale parfois supérieure, avec des informations dans le Moyen Infra-rouge, cependant leur résolution spatiale était toujours très inférieure (pixels de 20 m de côté). Le couple image THRS – analyse orientée-objet présente donc des résultats plus pertinents pour la cartographie de ces groupements végétaux, malgré les erreurs résiduelles.
Il a par exemple permis de préciser la localisation d’habitats intermédiaires encore non cartographiés. À ce titre, on peut citer les groupements mixtes roselières-magnocariçaies qui abritent certaines espèces floristiques rares et protégées, comme la Grande Douve (Ranunculus lingua), la Stellaire des marais (Stellaria palustris) ou encore le Peucedan à feuilles en lanières (Peucedanum lancifolium).
L’image Pléiades, du fait de sa période d’acquisition, a aussi permis d’identifier la Jussie (Ludwigia sp.), dont le pic de développement se situe au mois d’août, et dont l’expansion spatiale s’est très sensiblement accrue depuis 2009. Cependant, son identification est encore incomplète, là aussi à cause d’un décalage entre la perception humaine et l’identification par la machine. Cette dernière individualise très bien les herbiers denses, mais pose encore de nombreux problèmes de confusions avec les prairies. C’est pourquoi cette classe n’a pas été représentée ici (fig. 3).
Connaissance des habitats d’espèces
Ces résultats peuvent servir de fondement à un travail de cartographie des habitats potentiels de deux espèces d’oiseaux paludicoles. Ces deux espèces, le Butor étoilé (Botaurus stellaris) et le Phragmite aquatique (Acrocephalus paludicola), occupent l’une comme l’autre les zones humides, mais n’en font cependant pas le même usage. La première y niche principalement dans les phragmitaies denses, la seconde y fait une étape, principalement dans les formations d’hélophytes ouvertes ou à proximité des milieux dégagés, lors de sa migration post-nuptiale entre l’Europe de l’Est (Pologne, Biélorussie et Ukraine pour les principales populations) et l’Afrique sub-saharienne. Toutes deux sont considérées comme menacées, du moins à l’échelle européenne (listées à l’annexe I de la directive « Oiseaux » 79/409/CEE puis 2009/147/CE). Dans ce cadre, obtenir une carte fiable et précise de leurs habitats potentiels, sur la base des connaissances acquises dans le cadre des Plans Nationaux d’action (PNA) (Le Nevé et al., 2009 ; Hunault et Kerbiriou, 2007) devient un enjeu important. D’une part, cette carte permettrait de localiser les habitats qui peuvent jouer un rôle majeur dans le cycle biologique et la conservation de ces espèces à l’échelle du site (repos pour les migrateurs, alimentation, reproduction…). D’autre part, elle permettrait de cibler les campagnes d’inventaires ornithologiques, car tous les groupements ne sont pas exploités de la même manière par les oiseaux.
Une telle cartographie pourrait même à terme bénéficier d’un niveau de détail plus fin en terme d’habitat (au sein d’un même groupement). Il devrait par exemple être possible de différencier les roselières denses et celles émaillées de petits plans d’eau, ou encore les roselières jeunes de celles ayant tendance à l’atterrissement (présence ou non de Saules Salix sp.). Des contraintes matérielles (le manque de temps en particulier) nous ont cependant, pour l’instant, empêchés d’approfondir ces questions.
Conclusion
Atteignant un plus grand degré de précision spatiale, mais aussi de précision sémantique, la cartographie obtenue par analyse d’image orientée objet apporte des avancées notables dans la connaissance de la végétation des zones humides en général, du Pnr de Brière en particulier, et permet de compléter les travaux portant sur les habitats d’espèces.
Facilement reproductible de par son caractère automatique, cette méthode de cartographie ouvre des possibilités de suivis diachroniques des groupements végétaux à l’échelle de l’ensemble du marais. Elle complète ainsi efficacement les travaux d’inventaires botaniques de terrain, qui restent cependant indispensables pour des identifications précises, l’inventaire des plantes rares et protégées, mais aussi évidemment pour contrôler le travail de cartographie par télédétection et analyse d’image.
Les résultats obtenus peuvent être intégrés dans un Système d’Information Géographique (SIG), ce qui leur permet d’être utilisés dans d’autres travaux du Pnr. A titre d’exemple, le recensement des boisements au sein de la Grande Brière Mottière facilite leur gestion : lutte contre la fermeture du paysage, ou de façon plus pragmatique, exploitation du bois de chauffage. De façon plus spéculative, cette méthode de cartographie pourrait aussi permettre le recensement des mares et des haies (grâce à l’intervention de critères de forme), et ainsi contribuer à l’analyse de la structure du bocage, et du paysage associé (Burel et Baudry, 1999). Par ce biais, ces cartes faciliteraient les diagnostics effectués dans le cadre des programmes de Trame Verte et Bleue (TVB), le Pnr de Brière n’abritant pas uniquement des zones humides.